









































مقدمه
جیبرلین یکی از هورمونهای گیاهی است که نقش مهمی در رشد و نمو گیاهان بازی میکند. نام جیبرلینها به علت کشف تصادفی در ژاپن در قارچ آسکومیست انگل برنج به نام ژیبرلافوزیکوروژآ میباشد. این قارچ مسئول بیماری باکانه است که گیاهان مبتلا به این بیماری با طویل شدن قابل ملاحظه میانگرهها مشخص میشوند که در این صورت به آنها گیاهکهای سرکش میگفتند. تماما پژوهشهای اولیه درباره ژیبرلینها عملا از کارها کوروساوا فیزیولوژیست ژاپنی حاصل شده است. تعداد جیبرلینهای شناخته شده اکنون متجاوز از 100 عدد است.
 |
اندازهگیری جیبرلینها
برای جیبرلینها آزمونهای زیستی مخصوص تعیین شده است. در سال 1957 فاینی از اختصاصات جیبرلینها در برطرف کردن کوتاه قدی منشا وراثتی بعضی از واریتههای نخود و ذرت برای اندازهگیری این تنظیم کنندههای رشد استفاده کرده است. سایر آزمونها بر روی اثرات دیگر فیزیولوژیکی ژیبرلینها صورت میگیرد که در این آزمونها از افزایش طول محور زیر لپه کاهو ، تولید آلفا آمیلاز توسط یاختههای واجد آلورون ، میوه جو استفاده شده است.
خصوصیات شیمیایی جیبرلینها
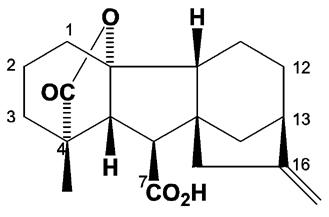
جیبرلینها بوسیله بوسیله پالک در سال 1965 به عنوان ترکیباتی با اسکلت انت جیبرلان شناخته شدند و در واقع دی ترپنهای حلقوی هستند که در نتیجه تراکم چهار واحد ایزوپرن بدست میآیند. جیبرلینها به شکل 19 و 20 اتم کربن ، منو ، دی و تری کربوکسیلیک غیر پیوسته وجود دارند. جیبرلینها بطور طبیعی در سه حالت یا شکل شیمیایی وجود دارند که دو نوع آنها از نظر شیمیایی مشخص شده است و سومی بطور فرضی معلوم گشته است. که عبارتند از:
- جیبرلینهای آزاد
- جیبرلینهای پیوسته
- جیبرلینهای محلول در آب
بیوسنتز جیبرلینها
جونز و فیلیس در سال 1966 نشان دادند که طرحهای اولیه برگی نسبت به مریستمهای راسی ، ژیبرلین بیشتر تولید میکنند. ریشهها به روش بسیار فعال این تنظیم کنندههای رشد را میسازند. جنین دانهها و میوهها نیز منبع خوب تولید جیبرلین هستند.
 |
انتقال جیبرلینها
بیشتر دادههای مربوط به انتقال جیبرلینها در گیاهان دانه دار به حرکت GA خارجی مربوط است و اطلاعات مربوط به حرکت GA درونی کاملا غیر مستقیم هستند. انتقال جیبرلین بطور غیر قطبی توسط آبکش و چوب صورت میگیرد. در نخود جیبرلینهای بکار برده شده به سرعت 5 سانتیمتر در ساعت حرکت میکند که این سرعت به سرعت متابولیتهای منتقل شده توسط آبکش مربوط است. جیبرلینها در شیره خام چند گیاه مو ، سیب و گوجه فرنگی یافت شده اند.
تشکیل مشتقات غیر فعال جیبرلینها
بتا هیدروکسیلاسیون
الف- هیدروکسیلاسیون جیبرلین ها در موقعیت 2 (β) به همراه کاهش محسوس فعالیت زیستی آنهاست. تبدیل GA1 به GA2 در دو گونه لوبیا (فازئولوس ولگاریس و فازئولوس کوکسینئوس) مشاهده شده است.
اشکال پیوسته جیبرلینها
گلوکزیل- استرهای جیبرلینهای مختلف شناخته شده است که در آنها گلوکز بوسیله پیوند بتا گلوکوزیدی به یک عامل هیدروکسیل جیبرلینها متصل است. این شکل جیبرلین بدون تردید از اشکال غیر فعال است زیرا نتوانستهاند هیدرولیز این مشتقات را در بافتها نشان دهند.
اثرات فیزیولوژیکی جیبرلینها
اثر بر روی رشد
جیبرلینها رشد طبیعی بعضی از گیاهان کوتاه قد را امکانپذیر میسازند همچنین رشد طولی گیاهان طبیعی مانند برنج ، کاهو ، گندم و خیار را تحریک میکنند. جیبرلینها اثری شگفت آوربر روی بلند شدن ساقه گیاهان طوقی دارند. فعالیت تقسیم یاختهای مریستم زیر راسی واریتههای نخود کوتاه قد و ذرت به جیبرلینها حساس بوده و بوسیله این هورمونها تحریک میشود. کشش یاختهای بیش از تقسیم یاختهای دلیل تحریک رشد بوسیله جیبرلین است. در واقع این هورمونها در گیاهان گندم یا جو که اشعه دیدهاند و در آنها تقسیم یاختهای کاملا متوقف شده هنوز موثر میباشند.
به علاوه بوسیله جیبرلیک اسید|جبرلیک اسید|GA3GA3 یک نوع همبستگی بین تحریک رشد محور زیر لپه کاهو و افزایش وزن خشک دیواره ملاحظه شده است. این رشد طولی با یک نوع افزایش قابل توجه دیکتیوزومها ، بوسیله یکنوع تکثیر آندوپلاسمی و افزایش پلیریبوزومهای یاختهها جلو میافتد. این تغییرات با سنتز و ترشح زیاد مواد سازنده دیواره توام است که شبیه تغییراتی است که توسط GA3 در یاختههای واجد آلورون گندم گیاه جو القا میشود. در یاختههای فوق این تغییرات قبل از افزایش سنتز و ترشح آلفا آمیلاز صورت میگیرد.
سنتز جدید هیدرولازها در یاختههای آلورون جو وابسته به GA
القای ساخت جدید آلفا آمیلاز بوسیله GA در لایههای آلورون پس از زمان تاخیری حدود 18 ساعت پس از عمل هورمون کاملا مشهود است. آلفا آمیلاز ظاهرا پس از ساخت از یاختههای آلورون آزاد میشود اما ریبونوکلئاز و بتا1و3 گلوکاناز ابتدا در یاختههای آلورون انباشته میشوند و سپس به سرعت آزاد شده یا ترشح میشوند. بطور کلی اگر قرار است سنتز جدید هیدرولازها ادامه یابد جیبرلین بطور مداوم حضور داشته باشد تا پاسخهای هورمون باقی بمانند.
ساخت RNA جدید برای القای سنتز جدید و آزاد شدن آلفا آمیلاز ممانعت میکند. بازدارندههای ساخت پروتئین از جمله سیکلو هگزیمید نیز از القای ساخت هیدرولازها توسط GA ممانعت میکنند. جالب است که اسید آبسیزیک که یک هورمون بازدارنده رشد است نیز از سنتز القا شده آلفا آمیلاز ممانعت میکند.
 |
سایر اثرات جیبرلین
جیبرلینها بخصوص GA7 و GA4 موجب تولید میوههای بدون دانه یا بکرباری در بعضی گونههای گیاهی میشود. همچنین اگر جیبرلیها به مقدار زیاد بکار رود یا به عنوان همکاری با سیتوکینین موجب رشد زیاد برگها میشود که غالبا سطح برگها به دو برابر طبیعی میرسد. جیبرلین در بیشتر موارد خفتگی دانهها را نیز برطرف میکند. بخصوص در مورد دانههایی که خفتگی حساس به نور دارند مانند کاهو که جیبرلین جانشین نور سرخ میشود و مانند نور سرخ خفتگی این گونه دانهها را برطرف میکند در این عمل معمولا اسید جیبرلیک به غلظت نسبتا قوی بکار برده میشود ( 3- 10 گرم در میلیلیتر).
جیبرلینها میتوانند بازدارندگی رویش دانه را که توسط اسید آبسیزیک در مورد جنینهای نخود حاصل شده برطرف نمایند و متقابلا اسید آبسیزیک میتواند اثر محرک جیبرلینها را تضعیف کند ولی در عین حال عمل آن عمومی نیست. عمل تضاد آبسیزیک که در چندین پدیده مشاهده میشود مربوط به یک نوع تشابه ساختاری به جیبرلین میتواند باشد. زیرا اسید آبسیزیک یک سزکوئیترپن است. اعمالی مانند خفتگی جوانهها با جیبرلین برطرف میشود. با کاربرد جیبرلینها ممکن است که این تنظیم کنندهها به عنوان آنتگونیتهای اسید آبسیزیک عمل کنند. جیبرلینها ورود به خفتگی جوانههای درختان و درختچهها را که بوسیله روزهای کوتاه القا میشود به تاخیر میاندازد.
آنتی جیبرلیک اسید
بعضی از کاهش دهندههای رشد که در برخی موارد عملی باغبانی برای بدست آوردن گیاهان پا کوتاه بکار میروند به نام آنتی جیبرلین معروفند. این ترکیبات سنتز جیبرلینها را متوقف میکنند یکی از انواع معروف این ترکیبات کلرور 2- کلرواتیل- تری متیل- آمونیوم است که در مسیر بیوسنتز جیبرلین سنتز کورن از ژرانیل- ژرانیول را متوقف میکند.